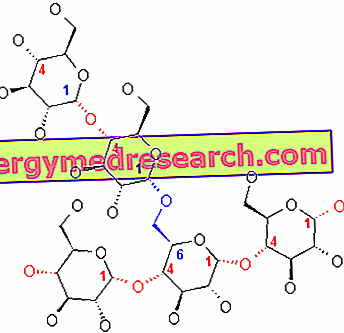
Glikogen to makrocząsteczka (masa cząsteczkowa około 400 milionów daltonów) α-glukozy, w której występują głównie wiązania i rozgałęzienia α-1, 4 glikozydowe w stosunku 1:10, ze względu na wiązania α-1, 6 glikozydowe.
Glikogen stanowi materiał rezerwowy i jest stale degradowany i odtwarzany; w całej masie komórek ciała znajduje się około 100 g glikogenu: większość znajduje się w wątrobie, gdzie jest ruchoma i dlatego może być wykorzystywana jako rezerwa dla innych narządów (glikogen w mięśniach nie jest ruchomy).
Enzymy, które katalizują degradację i syntezę glikogenu, wszystkie znajdują się w cytoplazmie, dlatego potrzebny jest system regulacji, który powoduje, że jedna droga jest nieaktywna, gdy druga jest aktywna: jeśli dostępna jest glukoza, ta druga jest przekształcana w glikogen (anabolizm), który jest rezerwą, odwrotnie, jeśli wymagana jest glukoza, to glikogen ulega degradacji (katabolizm).
Enzym zaangażowany głównie w degradację glikogenu jest fosforylazą glikogenu ; enzym ten jest w stanie rozszczepić wiązanie glikozydowe α-1, 4, stosując jako środek lityczny nieorganiczny ortofosforan: rozszczepienie zachodzi przez fosforolityczne i otrzymuje się 1-fosforan glukozy.
Przy pięciu lub sześciu jednostkach z punktu rozgałęzienia, enzym fosforylaza glikogenu nie jest już zdolny do działania, dlatego odłącza się od glikogenu i jest zastępowany przez enzym usuwający rozgałęzienia, który jest transferazą : w miejscu katalitycznym tego enzymu jest „histydyna, która umożliwia przeniesienie trzech jednostek sacharydowych do najbliższego łańcucha glikozydowego (histydyna atakuje pierwszy węgiel cząsteczki glukozy). Wspomniany enzym to glikozylotransferaza ; pod koniec działania tego enzymu tylko jedna jednostka glukozy pozostaje w łańcuchu bocznym, a pierwszy węgiel związany jest z szóstym węglem glukozy w głównym łańcuchu. Ostatnia jednostka glukozy łańcucha bocznego jest uwalniana przez działanie enzymu glikozydazy α-1, 6 (enzym ten jest drugą częścią enzymu usuwającego rozgałęzienia); biorąc pod uwagę, że gałęzie w glikogenie są w stosunku 1:10, z całkowitej degradacji makrocząsteczki otrzymujemy około 90% 1-fosforanu glukozy i około 10% glukozy.
Działanie wyżej wymienionych enzymów pozwala na eliminację łańcucha bocznego z cząsteczki glikogenu; aktywność tych enzymów można powtarzać, dopóki nie nastąpi całkowita degradacja łańcucha.
Rozważmy hepatocyt; glukoza (przyswajana przez dietę), kiedy wchodzi do komórki, jest przekształcana w glukozo-6-fosforan i w ten sposób aktywowana. Glukozo-6-fosforan, przez działanie fosfoglukomutazy, przekształca się w 1-fosforan glukozy: ten ostatni jest nie-bezpośrednim prekursorem biosyntezy; w biosyntezie stosuje się aktywowaną formę cukrów, która jest reprezentowana przez cukier związany z difosforanem: zwykle uridylodifosforan (UDP). 1-fosforan glukozy jest następnie przekształcany w glukozę UDP; ten metabolit znajduje się pod działaniem syntazy glikogenu, która jest zdolna do wiązania UDP-glukozy do nieredukującego końca rosnącego glikogenu: wydłużony glikogen otrzymuje się z jednostki glukozydowej i UDP. UDP jest przekształcany przez enzym nukleozydową fosfokinazę w UTP, który powraca do krążenia.
Degradacja glikogenu zachodzi poprzez działanie fosforylazy glikogenu, która uwalnia cząsteczkę glukozy i przekształca ją w 1-fosforan glukozy. Następnie fosfoglukomutaza przekształca 1-fosforan glukozy w 6-fosforan glukozy.
Glikogen jest syntetyzowany przede wszystkim w wątrobie i mięśniach: w organizmie znajduje się 1-1, 2 g glikogenu rozłożonego w całej masie mięśniowej.
Glikogen miocytu stanowi rezerwę energii tylko dla tej komórki, podczas gdy glikogen zawarty w wątrobie jest również rezerwą dla innych tkanek, to znaczy może być wysłany, jako glukoza, do innych komórek.
Glukozo-6-fosforan otrzymany w mięśniach z rozkładu glikogenu jest następnie przesyłany, w przypadku zapotrzebowania na energię, do glikolizy; w wątrobie glukozo-6-fosforan jest przekształcany w glukozę przez działanie fosfatazy 6-fosforanowej glukozy (enzymu charakterystycznego dla hepatocytów) i jest przenoszony do strumienia krążenia.
Zarówno syntaza glikogenu, jak i fosforylaza glikogenu działają na jednostki nieredukujące glikogenu, więc musi istnieć sygnał hormonalny, który nakazuje aktywację jednej ścieżki i blokowanie drugiej (lub odwrotnie).
W laboratorium możliwe było wydłużenie łańcucha glikogenu, wykorzystanie fosforylazy glikogenu i zastosowanie 1-fosforanu glukozy w bardzo wysokim stężeniu.
W komórkach fosforylaza glikogenowa tylko katalizuje reakcję degradacji, ponieważ stężenia metabolitów są takie, że przesuwają równowagę następującej reakcji, w prawo (tj. W kierunku degradacji glikogenu):

Spójrzmy na mechanizm działania fosforylazy glikogenu: tlen acetalowy (który działa jako pomost między jednostkami glukozy) wiąże się z wodorem fosforylu: półprodukt reakcyjny tworzy się przez karbokation (na glukozie, która jest kończyna), z którą wiąże się bardzo szybko fosforyl (Pi).
Fosforylaza glikogenu wymaga kofaktora, którym jest fosforan pirydoksalu (ta cząsteczka jest również kofaktorem dla aminotransferaz): ma protonowaną fosforyl tylko częściowo (fosforan pirydoksalu jest otoczony przez środowisko hydrofobowe, które uzasadnia obecność związanych z nim protonów), Fosforyl (Pi) jest w stanie dostarczyć proton do glikogenu, ponieważ taki fosforyl odzyskuje wówczas proton z częściowo protonowanej fosforylu fosforanu pirydoksalu. Prawdopodobieństwo, że przy fizjologicznym pH fosforyl traci proton i pozostaje całkowicie deprotonowane, jest bardzo niskie.
Zobaczmy teraz, jak działa fosfoglukomutaza. Enzym ten ma fosforylowaną resztę seryny w miejscu katalitycznym; seryna daje fosforyl do 1-fosforanu glukozy (w pozycji szóstej): w krótkim czasie tworzy się 1, 6-bisfosforan glukozy, a następnie sefosforyluje się przez przyjęcie fosforylu w pozycji pierwszej. Mutaza fosfoglucus może działać w obu kierunkach, tj. Przekształcać glukozo-1-fosforan w glukozo-6-fosforan lub odwrotnie; jeśli wytwarzany jest 6-fosforan glukozy, można go bezpośrednio wysłać do glikolizy do mięśni lub przekształcić w wątrobę w glukozę.
Enzym uridylofosfoglukotransferaza (lub pirofosforylaza glukozowa UDP) katalizuje reakcję przenoszenia 1-fosforanu glukozy na UTP przez atak fosforylowy.
Opisany właśnie enzym jest pirofosforylazą: ta nazwa wynika z faktu, że przeciwną reakcją do opisanej właśnie jest pirofosforylacja.
Uzyskana jak opisano glukoza UDP jest w stanie wydłużyć łańcuch glikogenu w jednostce monosacharydowej.
Możliwe jest rozwinięcie reakcji w kierunku tworzenia glukozy UDP przez wyeliminowanie produktu, który jest pirofosforanem; enzym pirofosfataza przekształca pirofosforan w dwie cząsteczki ortofosforanu (hydroliza bezwodnika) i dzięki temu utrzymuje stężenie pirofosforanu na tak niskim poziomie, że proces tworzenia glukozy UDP jest faworyzowany termodynamicznie.
Jak wspomniano, glukoza UDP, dzięki działaniu syntazy glikogenu, jest w stanie wydłużyć łańcuch glikogenu.
Konsekwencje (w stosunku 1:10) wynikają z faktu, że gdy łańcuch glikogenu składa się z 20-25 jednostek, interweniuje enzym rozgałęziający (mający histydynę w miejscu katalitycznym), zdolny do przeniesienia serii 7 -8 jednostek glikozydowych dalej od 5-6 jednostek: w ten sposób powstaje nowe rozgałęzienie.
Z powodów pochodzenia nerwowego lub jeśli energia jest potrzebna z powodu wysiłku fizycznego, adrenalina jest wydzielana z nadnerczy.
Docelowymi komórkami adrenaliny (i noradrenaliny) są komórki wątroby, mięśni i tkanki tłuszczowej (w tym ostatnim dochodzi do degradacji triglicerydów i krążenia kwasów tłuszczowych: w mitochondriach jest to zatem glukoza 6) -fosforan, który ma być wysłany do glikolizy, podczas gdy w adipocytach, 6-fosforan glukozy jest przekształcany w glukozę przez działanie enzymu fosfatazy glukozo-6-fosforanowej i eksportowany do tkanek).
Zobaczmy teraz sposób działania adrenaliny. Adrenalina wiąże się z receptorem umieszczonym na błonie komórkowej (miocytów i hepatocytów), co determinuje translację sygnału z zewnątrz do wnętrza komórki. Kinaza białkowa jest aktywowana, która działa jednocześnie na układy, które regulują syntezę i degradację glikogenu:
Syntaza glikogenu występuje w dwóch postaciach: postać defosforylowana (aktywna) i postać fosforylowana (nieaktywna); kinaza białkowa fosforyluje syntazę glikogenu i blokuje jego działanie.
Fosforylaza glikogenowa może występować w dwóch postaciach: w postaci aktywnej, w której obecna jest fosforylowana seryna, oraz w postaci nieaktywnej, w której seryna jest defosforylowana. Fosforylaza glikogenowa może być aktywowana przez enzym kinazę fosforylazy glikogenu . Kinaza fosforylazy glikogenu jest aktywna, jeśli jest fosforylowana i nieaktywna, jeśli jest defosforylowana; kinaza białkowa ma jako substrat kinazę fosforylazy glikogenowej, która jest zdolna do fosforylacji (a zatem aktywacji) tej ostatniej, która z kolei aktywuje fosforylazę glikogenu.
Gdy sygnał adrenaliny się skończy, efekt, jaki ma w komórce, musi również zakończyć się: enzymy fosfatazy interweniują zatem na gatunek białka.