Aby móc mówić kompleksowo o hemoglobinie (Hb), warto najpierw poradzić sobie z mioglobiną (Mb), która jest bardzo podobna do hemoglobiny, ale jest znacznie prostsza. Istnieją ścisłe związki pokrewieństwa między hemoglobiną i mioglobiną: oba są sprzężonymi białkami, a ich grupa prostetyczna (część niebiałkowa) jest grupą hemu .
Mioglobina jest białkiem kulistym składającym się z pojedynczego łańcucha około stu pięćdziesięciu aminokwasów (w zależności od organizmu), a jego masa cząsteczkowa wynosi około 18 Kd.
Jak wspomniano, jest on wyposażony w grupę hemową, która jest wstawiona w hydrofobową (lub lipofilową) część białka, składającą się z fałd, które można przypisać strukturom α-helisy białek włóknistych.
Mioglobina składa się głównie z segmentów α-helis, obecnych w liczbie ośmiu i składa się prawie wyłącznie z reszt niepolarnych (leucyna, walina, metionina i fenyloalanina), podczas gdy reszty polarne są praktycznie nieobecne (kwas asparaginowy, kwas glutaminowy, lizyna i arginina); jedynymi resztami polarnymi są dwie histydyny, które odgrywają zasadniczą rolę w ataku tlenu na grupę hemową.
Grupa hemowa jest grupą chromoforową (absorbuje w widzialnym) i jest grupą funkcyjną mioglobiny.
Zobacz także: hemoglobina glikowana - hemoglobina w moczu
Trochę chemii
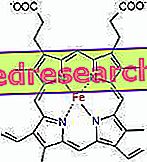
Związek między protoporfiryną a żelazem jest typowym wiązaniem tak zwanych związków koordynacyjnych, które są związkami chemicznymi, w których centralny atom (lub jon) tworzy wiązania z innymi substancjami chemicznymi w liczbie większej niż jego liczba utleniania (ładunek elektryczny). W przypadku hemu te wiązania są odwracalne i słabe.
Liczba koordynacyjna (liczba wiązań koordynacyjnych) żelaza wynosi sześć: wokół żelaza może być sześć cząsteczek, które dzielą elektrony wiążące.
Aby utworzyć związek koordynacyjny, wymaga dwóch orbitali o prawidłowej orientacji: jednej zdolnej do „kupowania” elektronów, a drugiej zdolnej do oddania ich
W hemie żelazo tworzy cztery płaskie wiązania z czterema atomami azotu, które znajdują się w środku pierścienia protoporfirynowego i piąte wiązanie z bliższym azotem histydyny; żelazo ma szóste wolne ogniwo koordynacyjne i może wiązać się z tlenem.
Kiedy żelazo jest w postaci wolnego jonu, jego orbitale typu d mają taką samą energię; w mioglobinie jon żelaza wiąże się z protoporfiryną i histydyną: gatunki te magnetycznie zakłócają orbitale żelaza; zakres zaburzeń będzie różny dla różnych orbitali w zależności od ich orientacji przestrzennej i przeszkadzających gatunków. Ponieważ całkowita energia orbitali musi powrócić do stałej, zaburzenie powoduje energetyczną separację pomiędzy różnymi orbitałami: energia pozyskana przez niektóre orbitale jest równoważna energii utraconej przez inne.
Jeśli separacja zachodząca między orbitalami nie jest zbyt duża, preferowany jest układ elektroniczny o wysokim spinie: elektrony wiążące próbują układać się równolegle na kilku możliwych poziomach (maksymalna wielokrotność); jeśli z drugiej strony zaburzenie jest bardzo silne i istnieje duża separacja między orbitałami, wygodniejsze może być sparowanie elektronów wiążących w niższych orbitali energetycznych (niskie obroty).
Gdy żelazo wiąże się z tlenem, cząsteczka przyjmuje niską dyspozycję spinową, podczas gdy żelazo ma szóste wolne połączenie koordynacyjne, cząsteczka ma wysokie obroty.
Dzięki tej różnicy w spinie, dzięki analizie spektralnej mioglobiny, jesteśmy w stanie zrozumieć, czy istnieje związany tlen (MbO2) czy nie (Mb).
Mioglobina jest białkiem typowym dla mięśni (ale nie tylko w mięśniach).
Mioglobina jest ekstrahowana z kaszalotu, w którym występuje w dużych ilościach, a następnie oczyszczana.
Walenie mają oddychanie podobne do ludzkiego: płuca muszą wchłaniać powietrze przez proces oddechowy; kaszalot musi nosić jak najwięcej tlenu w mięśniach, które są w stanie gromadzić tlen, wiążąc go z obecną w nich mioglobiną; tlen jest następnie uwalniany powoli, gdy walenie jest zanurzone, ponieważ jego metabolizm wymaga tlenu: im większa ilość tlenu, którą kaszalot jest w stanie wchłonąć i tym więcej tlenu jest dostępne podczas nurkowania.

Aby mięso było mniej lub bardziej czerwone, zawartość hemoprotein jest czerwona (mięso to jest hemem).
Hemoglobina ma wiele analogii strukturalnych z mioglobiną i jest zdolna do odwracalnego wiązania tlenu cząsteczkowego; ale chociaż mioglobina jest ograniczona do mięśni i ogólnie tkanek obwodowych, hemoglobina znajduje się w erytrocytach lub czerwonych krwinkach (są to pseudokomórki, to znaczy nie są prawdziwymi komórkami), które stanowią 40% krwi.
Hemoglobina jest tetrametrem, to znaczy składa się z czterech łańcuchów polipeptydowych, każdy z grupą hemu i dwa po dwóch (u człowieka istnieją dwa łańcuchy alfa i dwa łańcuchy beta).
Główną funkcją hemoglobiny jest transport tlenu ; Inną funkcją krwi, w którą zaangażowana jest hemoglobina, jest transport substancji do tkanek.
Na drodze od płuc (bogatych w tlen) do tkanek hemoglobina przenosi tlen (jednocześnie inne substancje docierają do tkanek), podczas gdy na odwrotnej ścieżce przenosi ona ze sobą odpady zebrane z tkanek, zwłaszcza dwutlenek węgla wytwarzany w metabolizmie.
W rozwoju człowieka istnieją geny, które są wyrażane tylko przez pewien okres czasu; z tego powodu mamy różne hemoglobiny: płodową, embrionalną dorosłego człowieka.
Łańcuchy tworzące te różne hemoglobiny mają różne struktury, ale z podobieństwami funkcja, którą pełnią, jest mniej więcej taka sama.
Wyjaśnienie obecności kilku różnych łańcuchów jest następujące: podczas ewolucyjnego procesu rozwoju organizmów hemoglobina również ewoluowała, specjalizując się w transporcie tlenu z obszarów bogatych w obszary ubogie. Na początku łańcucha ewolucyjnego hemoglobina zawierała tlen w małych organizmach; w trakcie ewolucji organizmy osiągnęły większe wymiary, dlatego hemoglobina zmieniła się, aby móc transportować tlen do obszarów dalej od punktu, w którym była bogata; aby to zrobić, nowe struktury łańcuchów tworzących hemoglobinę zostały skodyfikowane w trakcie procesu ewolucyjnego.
Mioglobina wiąże tlen nawet przy niewielkim ciśnieniu; w tkankach obwodowych występuje ciśnienie (PO2) wynoszące około 30 mmHg: przy tym ciśnieniu mioglobina nie uwalnia tlenu, więc byłaby nieskuteczna jako nośnik tlenu. Z drugiej strony hemoglobina ma bardziej elastyczne zachowanie: wiąże tlen pod wysokim ciśnieniem i uwalnia go, gdy ciśnienie spada.
Gdy białko jest funkcjonalnie aktywne, może nieznacznie zmienić swój kształt; na przykład natlenowana mioglobina ma inną postać niż nietlenowana mioglobina i ta mutacja nie wpływa na sąsiednie.
Dyskurs jest inny w przypadku powiązanych białek, takich jak hemoglobina: gdy łańcuch jest natleniony, indukuje się zmianę jego kształtu, ale ta modyfikacja jest trójwymiarowa, dlatego wpływają również na inne łańcuchy tetrametru. Fakt, że łańcuchy są ze sobą powiązane, pozwala sądzić, że modyfikacja jednego oddziałuje na inne sąsiednie, nawet jeśli w innym stopniu; kiedy łańcuch jest natleniony, inne łańcuchy tetrametru przybierają „mniej wrogie nastawienie” wobec tlenu: trudność, z jaką łańcuch oksygenuje, zmniejsza się, gdy łańcuchy przy nim natleniają się. Ten sam argument dotyczy deoksygenacji.
Czwartorzędowa struktura deoksyhemoglobiny nazywana jest formą T (czas), podczas gdy struktura oksyhemoglobiny nazywana jest formą R (uwalniana); w stanie rozciągniętym istnieje szereg raczej silnych oddziaływań elektrostatycznych między aminokwasami kwasowymi i aminokwasami zasadowymi, które prowadzą do sztywnej struktury deoksyhemoglobiny (dlatego „forma napięta”), podczas gdy gdy tlen jest związany, istota tych zmniejszają się interakcje (dlatego „uwolniona forma”). Ponadto, przy braku tlenu, ładunek histydynowy (patrz struktura) jest stabilizowany przez przeciwny ładunek kwasu asparaginowego, podczas gdy w obecności tlenu białko ma tendencję do utraty protonu; Wszystko to oznacza, że utlenowana hemoglobina jest silniejszym kwasem niż odtleniona hemoglobia: efekt bohr .
W zależności od pH grupa hemowa z łatwością wiąże się z tlenem: w środowisku kwaśnym hemoglobina łatwiej uwalnia tlen (forma napięta jest stabilna), podczas gdy w środowisku podstawowym wiązanie z tlenem jest silniejszy.

Każda hemoglobina uwalnia 0, 7 protonów na przychodzący mol tlenu (O2).
Efekt Bohra pozwala hemoglobinie poprawić jej zdolność do przenoszenia tlenu.
Hemoglobina, która powoduje przejście z płuc do tkanek, musi się równoważyć w zależności od ciśnienia, pH i temperatury.
Widzimy wpływ temperatury .
Temperatura w pęcherzykach płucnych jest o 1-1, 5 ° C niższa niż temperatura zewnętrzna, podczas gdy w mięśniach temperatura wynosi około 36, 5-37 ° C; wraz ze wzrostem temperatury zmniejsza się współczynnik nasycenia (z tym samym ciśnieniem): dzieje się tak, ponieważ zwiększa się energia kinetyczna i sprzyja dysocjacji.

Istnieją inne czynniki, które mogą wpływać na zdolność hemoglobiny do wiązania się z tlenem, z których jednym jest stężenie 2, 3 bisfosfogliceryny .
2, 3 bis-fosfogliceryd jest czynnikiem metabolicznym obecnym w erytrocytach w stężeniu 4-5 mM (w żadnej innej części ciała nie występuje w tak wysokim stężeniu).
Przy fizjologicznym pH 2, 3 bisfosfoglicerynian jest deprotonowany i zawiera pięć ładunków ujemnych; przechodzi w klin między dwoma łańcuchami beta hemoglobiny, ponieważ łańcuchy te mają wysokie stężenie ładunków dodatnich. Oddziaływania elektrostatyczne między łańcuchami beta i łańcuchami bisfosfoglicerynianowymi nadają systemowi pewną sztywność: uzyskuje się strukturę napiętą, która ma małe powinowactwo do tlenu; podczas natleniania wypędza się 2, 3 bis-fosfoglicerynian.
W erytrocytach znajduje się specjalny aparat, który przekształca 1, 3-bisfosfogliceryd (wytwarzany przez metabolizm) w 2, 3 bis-fosfogliceryd, tak że osiąga stężenie 4-5 mM, a zatem hemoglobina może wymieniać l tlen w tkankach.
Hemoglobina docierająca do tkanki jest w stanie uwolnionym (związanym z tlenem), ale w pobliżu tkanki jest karboksylowana i przechodzi w stan rozciągnięty: białko w tym stanie ma mniejszą tendencję do wiązania się z tlenem, w porównaniu do stanu uwolnionego, dlatego hemoglobina uwalnia tlen do tkanki; ponadto, w wyniku reakcji pomiędzy wodą i dwutlenkiem węgla, następuje wytwarzanie jonów H +, a zatem dalszy tlen dzięki efektowi bohr.
Dwutlenek węgla dyfunduje do erytrocytów przechodzących przez błonę plazmatyczną; biorąc pod uwagę, że erytrocyty stanowią około 40% krwi, powinniśmy się spodziewać, że tylko 40% dwutlenku węgla, który dyfunduje z tkanek, wchodzi do nich, w rzeczywistości 90% dwutlenku węgla dostaje się do krwinek czerwonych, ponieważ zawierają enzym, który przekształca dwutlenek węgla w kwasie węglowym, wynika z tego, że stacjonarne stężenie dwutlenku węgla w erytrocytach jest niskie i dlatego szybkość wejścia jest wysoka.

Kolejne zjawisko, które występuje, gdy erytrocyt dociera do tkanki, jest następujące: przez gradient, HCO3- (pochodna dwutlenku węgla) wychodzi z erytrocytów i, aby zrównoważyć wyjście ładunku ujemnego, mamy wprowadzanie chlorków, które określa wzrost ciśnienia osmotycznego: aby zrównoważyć tę zmienność, występuje również wejście wody, która powoduje puchnięcie erytrocytów (efekt HAMBURGERA). Przeciwne zjawisko występuje, gdy erytrocyt dociera do pęcherzyków płucnych: występuje deflacja erytrocytów (efekt HALDANE). Zatem żylne erytrocyty (bezpośrednio do płuc) są bardziej okrągłe niż tętnicze.